 用單個錐形光纖植入物進行深度分辨光纖光度測定
用單個錐形光纖植入物進行深度分辨光纖光度測定
用單個錐形光纖植入物進行深度分辨光纖光度測定
(轉譯自文獻Depth-resolved fiber photometry with a single tapered optical fiber implant)
活體熒光檢測可用于記錄和研究自由運動動物腦深部遺傳定義的神經群的功能信號。例如,纖維光度法通過監測特定細胞類型神經活動時熒光隨時間變化來實現。這些方法推動了基于光子學和光電子平臺技術以及使用多路復用技術記錄多個亞種群活動方法的發展。通常情況下,光纖測量方案依賴于扁平切割光纖進行刺激和收集熒光2-9,11 - 19。
然而,由于組織散射和吸收效應,扁平切割光纖的可訪問記錄深度僅限于光纖尖端附近,這與探針的幾何形狀相結合,決定了熒光激發和收集效率20,21。簡單的幾何計算表明,扁平切割光纖收集的信號量隨著與光纖面距離的增加而急劇減少。此外,重新配置收集幾何形狀以達到多個區域是不可能的,因為改變光收集場需要重新定位光纖。此外,扁平切割光纖的幾何形狀嚴重損害組織,在大腦中,甚至在植入后很長一段時間內,也會誘導裝置周圍的神經膠質激活22,23。盡管如此,平劈光纖被廣泛用于評估腦深部區的神經活動3,11-19。
在這里,我們提出了一種克服這些限制的方法:我們利用TF中光傳播的模態特性在錐度的大光學活性區域上構造光收集模式并進入更深的細胞。除了比扁平切割光纖22具有更小的侵入性外,TF探針還具有獨特的光收集特征,包括:(i)沿光纖軸在高達2mm的組織上具有均勻的界面,(ii)通過分時多路復用沿錐度進行多點收集的能力,以及(iii)通過微結構光纖錐度的非平面表面來設計任意收集體積的能力。
下面,我們量化了錐形光纖的三維(3D)光采集區域,發現錐形光纖在大區域(如小鼠的大腦皮質和紋狀體)均勻地收集熒光。當與大面積光傳輸相結合時22,24,這導致在有源光學表面相似的照明功率密度下,錐形光纖比扁平切割光纖的信號采集更高。這是因為大面積的錐形光纖可以提供更多的總照明功率,即更多的光子,同時將電池暴露在中等的功率密度下。我們的研究表明,通過利用選擇性光傳遞和收集,轉錄因子能夠在自由運動的動物中對功能性熒光信號進行多點探測,包括沿著纖維錐度動態記錄來自多個腦區的信號。我們通過在自由運動的小鼠中使用單個錐形光纖完成獎勵收集任務,快速掃描興奮光并同時監測背側和腹側紋狀體的多巴胺瞬變,證明了這種實驗的可行性。
最后,我們將控制光沿錐度傳播的模態效應與金屬涂層錐形光纖表面的微觀和納米結構相結合,從而設計了收集體積25,26。我們將收集體積限制在錐度表面的一個角部分,這樣,光學窗口位于沿著錐形光纖界面的特定深度,只有很少的細胞體。這種方法與光學窗口的選擇性光傳輸相結合,提供了具有高度空間選擇性的深度細胞體積的雙向接口。
結果
錐形光纖的光收集特性
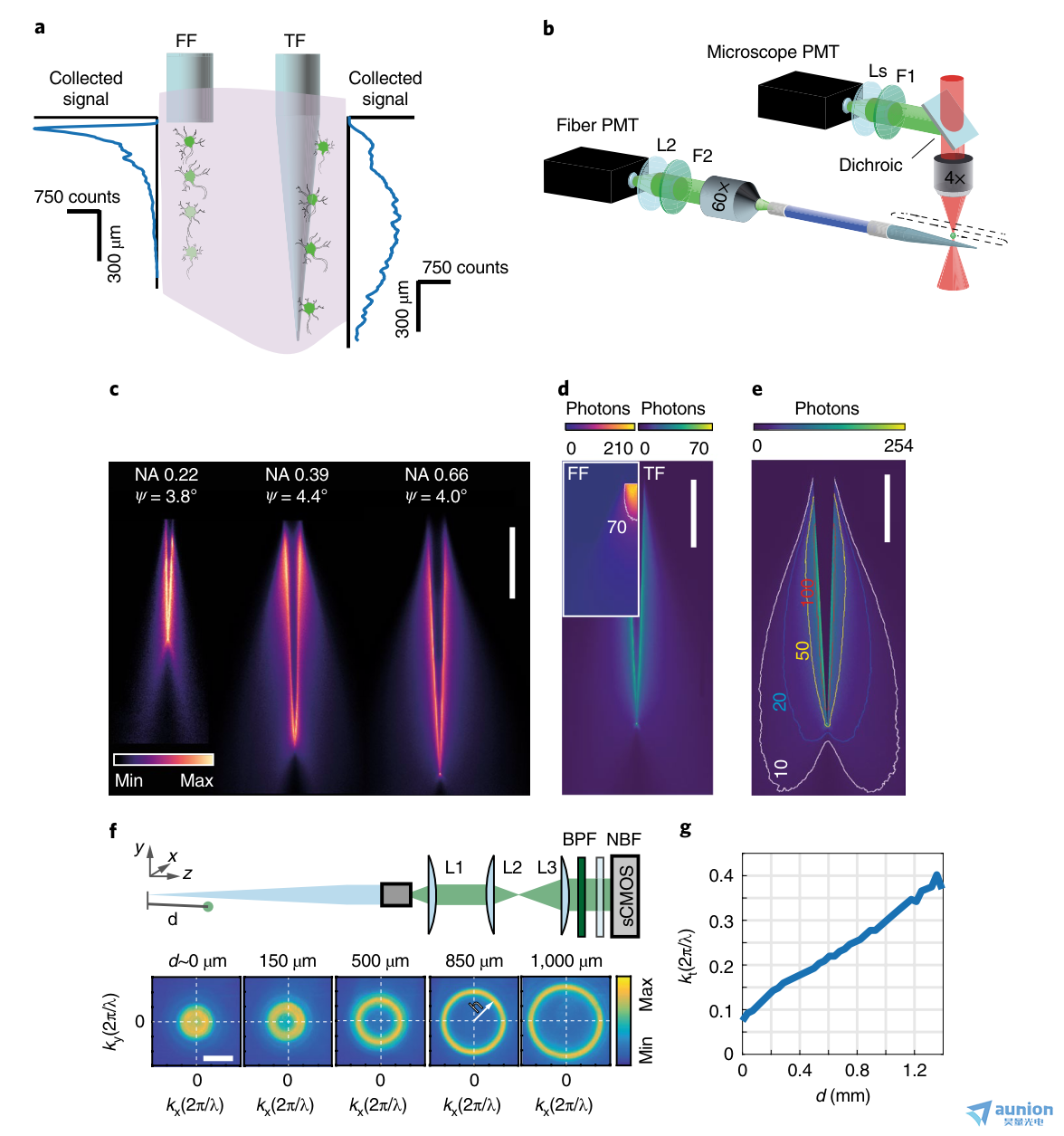
圖1 |錐形光纖的光收集。a,腦組織中扁平切割光纖(FF)和錐形光纖(TF)的光采集示意圖。實驗收集概況旁邊的纖維。b,對錐形光纖的光采集場成像的光學設置。在pbs -熒光素滴中,一個圍繞錐形光纖的雙光子激發點被掃描。產生的熒光可通過未脫膜的PMT(顯微鏡PMT)和補片光纖遠端的光纖PMT檢測。Ls,透鏡系統;F1和F2,帶通熒光濾波器;L2,鏡頭。c,在PBS-熒光素溶液中,隨著NAs的增加錐形光纖的典型ξT(x,y)集合字段(每個字段歸一化到其zui大值);比例尺,500μm。d,比較在pbs -熒光素溶液中掃描的雙光子熒光光斑采集的光子數(像素停留時間,3.2μs),內嵌扁平切割光纖與NA = 0.66, ψ = ~4°的錐形光纖;FF圖中的等值線顯示錐形光纖收集到的zui大光子數。比例尺,500μm。e, NA-0.66 錐形光纖在pbs -熒光素溶液中的光子收集的等距線(頂部色條,每個像素的光子數;停留時間,3.2μs);等值線在10、20、50和100光子處繪制。比例尺,500μm。f,上,遠場成像系統示意圖。L1、L2、L3,成像鏡;BPF,帶通濾波器;NBF,近紅外阻斷濾波器;sCOMS,科學互補金屬氧化物半導體。底部,纖維輸出小關節的遠場圖像顯示,當光源沿著錐形光纖移動時,直徑增加的環。比例尺,0.3 2π/λ。g, 錐形光纖在距離錐尖d處采集的點狀光源熒光的橫向矢量分量kt。a-d的實驗重復了至少10次,得到了相似的結果。
我們在準透明的熒光溶液中表征了錐形光纖的光聚集特性(圖1)。我們在浸泡錐形的pbs熒光素(30μM)液滴中實現了一個雙光子掃描系統,以產生局限的熒光斑,就像各向同性的點狀源一樣(圖1b)。光柵掃描錐度周圍光斑時產生的熒光由與掃描頭同步的兩個光電倍增管(PMT)收集:(i)顯微鏡PMT,放置在標準的非脫封,外熒光路徑,和(ii)光纖PMT,置于連接的光纖貼片的遠端至錐形光纖20、21(圖1b)。用顯微鏡PMT得到的參考圖像對視場中雙光子激發效率的輕微不均勻性進行校正后,來自光纖PMT的信號報告了錐形光纖的熒光光采集場,定義為ξT(x,y)。測量了不同數值孔徑(NAs)和芯徑,但錐度角(ψ)近似為~4°的光纖的集合場ξT(x,y)(圖1c)。我們發現沿錐度的光敏區域,即收集長度L,隨著光纖NA的增大和ψ的減小而增大(補充圖1a)。因此,錐形光纖的采集長度是可以定制的通過修改光纖NA和錐度角ψ,從幾百微米提高到約2 mm。這一發現揭示了錐形光纖和扁平切割光纖的收集特性的重要差異,因為對于扁平切割,收集深度基本上不依賴于NA21。
我們比較了錐形光纖和扁平切割的采集字段,NA分別為0.66(圖1d)和0.39(補充圖1b)。錐形光纖的光學主動表面沿波導軸線延伸,導致沿錐度方向相對均勻的收集。從集合字段ξF(x,y)中可以看出,扁平切割光纖在端面附近采集到較高的信號強度。相反,錐形光纖的收集效率曲線在錐度面附近達到一個較低的zui大值,并遵循在尖端增寬的兩葉形狀(圖1e和補充圖1c、d和2)如圖ξ(x,y,z)區域所示(補充圖1d),被采集信號圍繞錐度軸完全對稱。
這是因為錐形光纖表面通過增加波導直徑27的橫向傳播分量kt的模態子集與周圍環境進行光學界面。因此,由光纖的直部分所支持的全部傳播模式逐漸沿錐形填充,導致錐形光纖軸的均勻收集。相反,扁平切割光纖的所有傳播模式都在纖維面耦合。
為了更好地表征錐度采集光的物理特性,我們從靠近錐度表面的點樣點對熒光進行雙光子激發時,對所采集光的遠場進行成像(圖1f)。我們發現不同的模態子集在特定的錐度直徑下被填充(圖1f,g),因為相機上的圖像是一個環,它的半徑隨著熒光源和錐度尖端之間的距離的函數而增加。環半徑h是直接測量與進入纖維的導模相關的波矢量的橫向分量kt 27,28。因此,光線從錐體的不同截面進入,受到不同的引導模式子集的引導,在相機上產生不同直徑的環,從而建立了h與熒光信號沿錐體的位置之間的相關性。
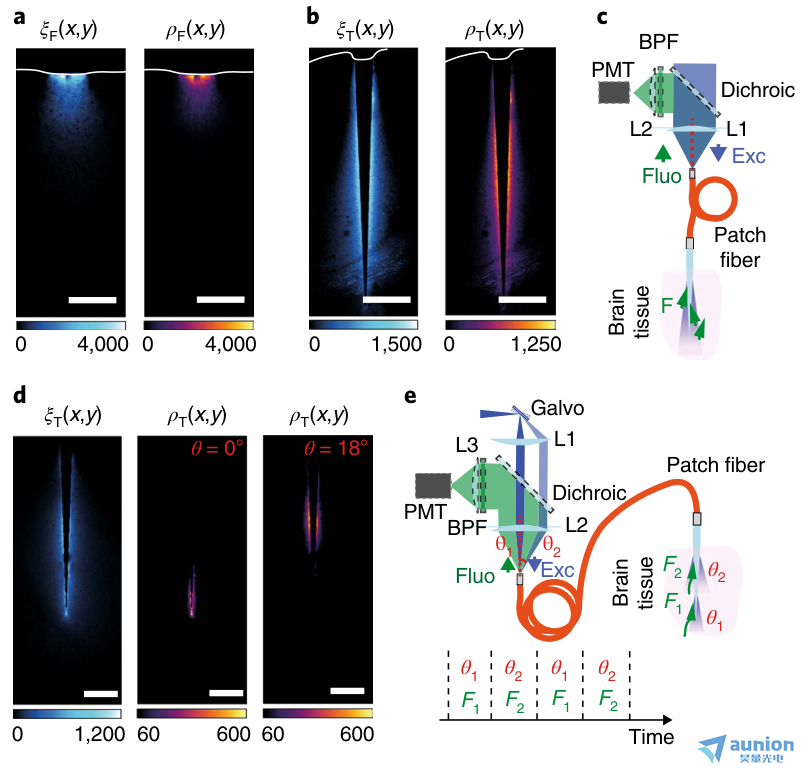
圖2 |可重構的錐形光纖光收集。a,與熒光素均勻染色腦片皮質接觸的0.66 NA 扁平切割光纖的光采集場ξ(x,y)(左)和光度效率場ρ(x,y)(右)。b,與a一樣,將0.66-NA 錐形光纖插入經熒光素均勻染色的腦切片中。c,使用全NA照明和藍色激光刺激并收集大大腦區域熒光的系統示意圖。收集的光在貼片光纖中反向傳播,并通過一個二色鏡將其導向PMT與藍光區分開來。L1和L2,晶狀體;BPF,帶通濾波器;Fluo,熒光信號;Exc,激發光。d,定點照明將采樣體積限制在錐形光纖的子區域。左,集光域ξT(x,y);中心,在光纖尖端選擇性照明得到的光度效率場ρT(x,y);右,ρT(x,y)視場是在較寬錐度直徑下選擇性照明獲得的。e,提出的多站點光度測量系統的原理圖,該系統使用時分復用配置的PMT探測器。藍色激光束以增加的輸入角(θ1, θ2)射入光纖貼片線。低角度注入時,激光在錐尖處耦合,產生熒光信號F1;相反,當在θ2處注入時,激光在較大錐度直徑下耦合,產生熒光信號F2。熒光由PMT檢測,其輸出信號與光注入刺激同步。該熒光信號根據其時間戳歸屬于相應的區域。a、b、d實驗重復三次,結果相似。
在大而深的區域統一收集
為了證明在存在散射和吸收的情況下,錐形光纖可以在大的和深部腦區獲得均勻的采集,我們測量了均勻熒光染色的腦片上扁平切割光纖和錐形光纖的熒光采集場ξ(x,y)和熒光激發場β(x,y)。結合這些場得到了光度測量效率場ρ(x,y),它描述了熒光信號對激發光強度的依賴性20,21,從而給出了采樣組織體積的詳盡幾何信息。我們比較了匹配NA和內核大小的扁平切割光纖和錐形光纖的采集和光度視野。如圖2a所示,插入到皮層表面的扁平切割光纖域ξF(x,y)和域ρF(x,y),扁平切割光纖有效地與皮層的淺層連接;然而,他們只提取了距離透鏡面300 μm以外的信息。相反,錐形光纖的界面更均勻,錐體的光學活性區域周圍有腦組織(圖2b)。利用ξ(x,y)的對稱性,我們計算了由波導采樣的體積作為采集信號的函數(補充圖3),并確定了產生給定比例的總采集信號的組織體積。我們發現錐形光纖的體積比扁平切割光纖大(補充圖4)。這一特性可以在使用全錐度表面來激發和收集信號的實驗中加以利用(圖2c)。
沿錐度可重新配置多站點收集
使用位點選擇性光傳輸和模分解復用策略,錐形光纖的收集量可以沿著錐度在多個位置之間動態切換22,27,28。為了定義可尋址的體積幾何配置,我們獲得了一個插入到熒光素染色腦片上的0.66-NA 錐形光纖的ξT(x,y)集合域(圖2d)。使用基于振鏡的快速掃描系統(圖2e),我們通過增加kt激發模態子集將激光注入錐形光纖,從而將照明體積限制在可通過改變光輸入角度22、27、28沿錐度部分逐漸移動的有限區域(補充圖5a、b)。由于熒光只在有限的被照射組織中產生(補充圖5a、b),錐形光纖可以動態地檢查一個功能區的多個位點。作為原理證明,我們結合ξT(x,y)和β(x,y),測量了由選址照明產生的光度測量效率場ρT(x,y)。ρT(x,y)在可從光線注入角度推斷的有限區域內zui大(圖2d)。利用這一特性,熒光信號可以歸因于使用時分復用被照亮的大腦區域(圖2e)。這是通過增加輸入角(θ1, θ2)將激光發射到光纖補片線來激發沿錐度在限制位置耦合的不同模態子集來實現的。每個照明位置產生的熒光(分別為F1、F2)由錐度采集,在光纖補片線中反向傳播,由二色鏡識別,zui后由PMT檢測,PMT輸出信號與光注入刺激同步(圖2e)。
為了證明這種方法對可能由動物運動引起的模態混合有彈性22,我們在pbs -熒光素浴中進行深度分辨光度測量時,在手動搖動光纖貼片的同時監測了遠場模式。記錄到的強度波動<1%,遠場環直徑和厚度變化<0.8%(對于未受擾動的纖維;補充圖5c、d和補充視頻1、2)。
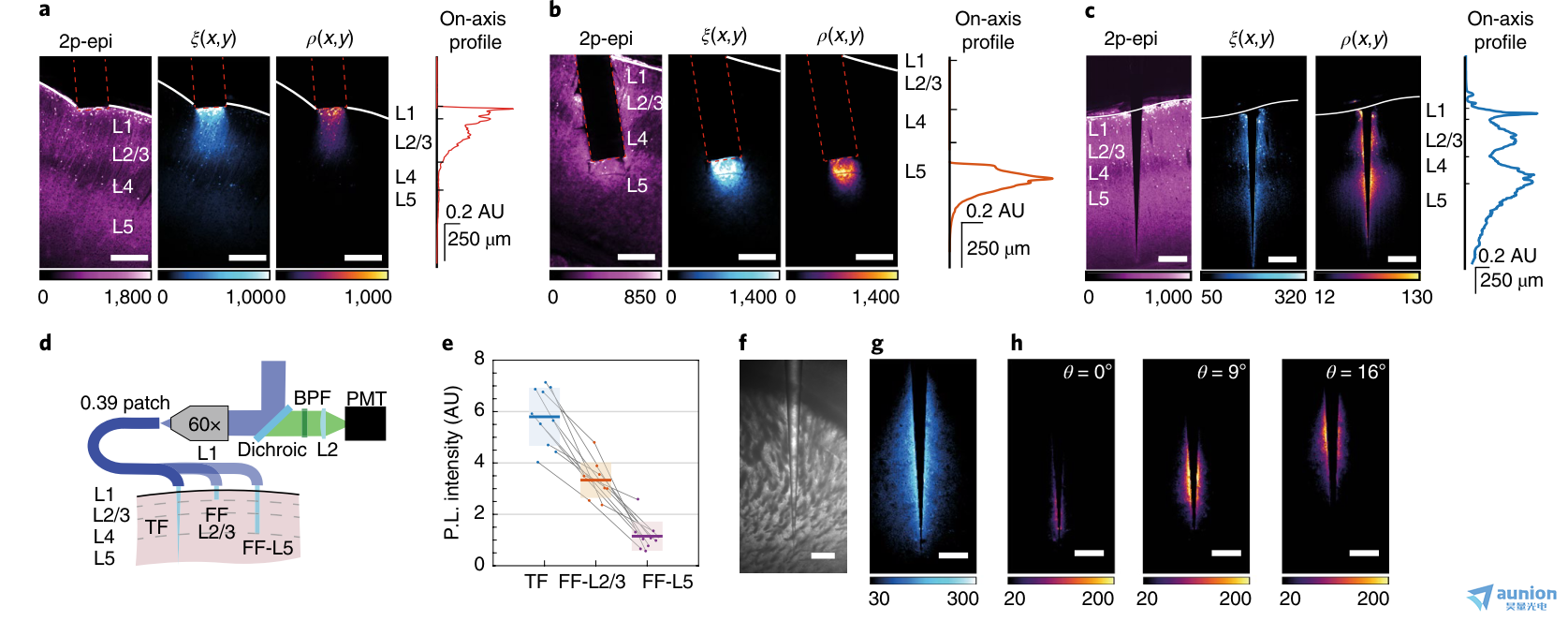
圖3 |基因染色的神經群增強的光度測定。a,皮質表面0.39-NA 扁平切割光纖的光采集;從左到右:雙光子表觀熒光(2p-epi)圖像,ξ(x,y)場,ρ(x,y)場,軸上采集輪廓ρ(x,y)。b, a為NA = 0.39 扁平切割光纖,接近L5。c, a,b表示一個在大腦皮層插入的0.39 NA的錐形光纖。比例尺(a?c), 250μm。d,三種實驗配置的光度測量系統示意圖:一個錐形光纖插入整個皮質,一個扁平切割光纖插入L2/3,一個扁平切割光纖插入至L5。e, Thy1-ChR2-EYFP小鼠腦片的熒光信號強度(n = 10),在大腦皮層插入0.39 NA 錐形光纖(ψ = 4°)(藍色),在L2/3插入0.39- NA 扁平切割光纖(橙色),在L5插入0.39- NA 扁平切割光纖(紫色)來刺激和檢測熒光。調整激光功率以獲得相似的光活性區域的功率密度(0.1 mW mm-2)。陰影區域表示平均值上的標準誤差。灰線連接在同一實驗中從同一腦片獲得的數據。采用雙側Student t檢驗進行統計學分析,顯著性α = 0.001。f, 錐形光纖插入固定腦片紋狀體的亮視野圖像(Thy1-ChR2-EYFP小鼠)。g, f. h中錐形光纖的光采集域ξT(x,y),將ξT(x,y)域與位點選擇性傳遞光相結合,產生可重構的紋狀體子區域多位點光采集效率域ρT(x,y)。比例尺(f?h), 250μm。在a-c、g、h重復實驗3次,結果相似。
增強熒光法在基因染色的神經群體
對于錐形光纖,我們使用了0.39-NA 錐形光纖(ψ = ~4°)和扁平切割光纖來刺激和檢測Thy1-ChR2-eYFP小鼠固定腦片不同皮質層的熒光,其中EYFP僅限于L2/3和L5(圖3a-c)。我們測量了三種實驗配置的ξ(x,y)、β(x,y)和ρ(x,y)場:靠近淺層的FF(圖3a)、插入L5層的FF(圖3b)和穿過皮質范圍的錐形光纖(圖3c)。正如預期的那樣,錐形光纖刺激并收集了L2/3和L5層的熒光,而扁平切割光纖在靠近關節突的一個有限區域內募集信號,需要重新定位以處理這兩個區域。此外,光纖纖維鈍的幾何輪廓阻礙了光纖的插入,因為當光纖穿過切片時,移位的組織仍然在關節突的前面。
我們比較了三種實驗配置產生的絕對信號水平(圖3d),通過調制激光功率來補償錐形光纖的較大光學活性區域,并在每個光學表面提供相同的平均功率密度(~0.1 mW mm-2)。在這些條件下,錐形光纖在兩個深度都相對于扁平切割光纖產生了更大的熒光信號(圖3e),這可以解釋為在光收集和光傳輸中分模解復用的綜合作用。在保持中等功率密度的情況下,模式分復用將較高的總照明功率分布在較寬的表面22,27。隨著更多的光子被釋放到組織中,更多的神經元參與到收集信號中,更多的熒光被產生和檢測到,這與之前的研究結果一致,在較低的輸出功率下錐形光纖比扁平切割光纖更能引起光遺傳激活22。重要的是,由于光漂白依賴于每個熒光團的光暴露,當全部的光活性區域被吸收時,錐形光纖在更大體積的組織上的光分布允許產生更多的熒光而不增加光漂白。
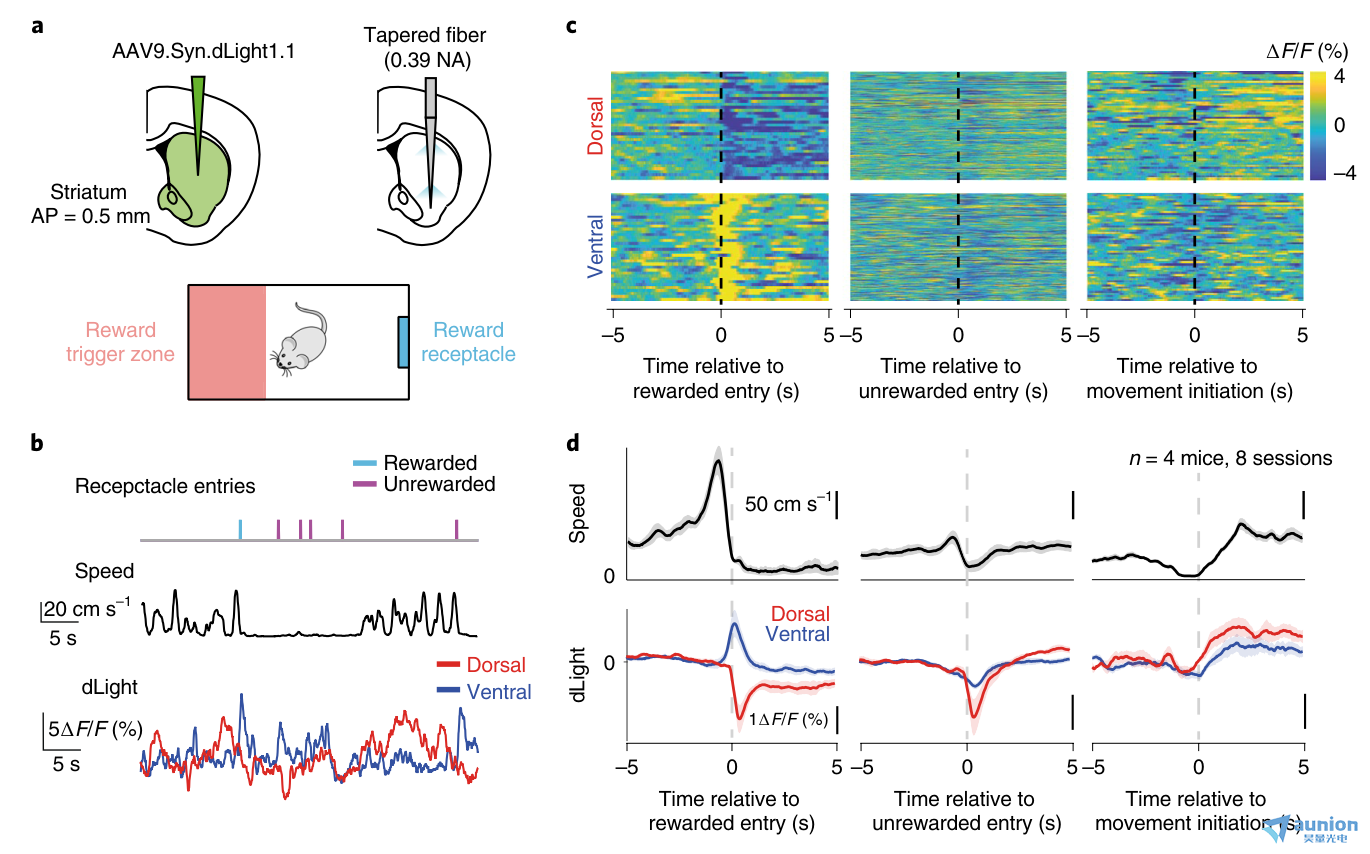
圖4 |體內多點光度法揭示了多巴胺對背側紋狀體和腹側紋狀體運動和獎勵的不同反應。a,頂部,用于活體光纖光度測定的手術部位和兩個部位錐形光纖照明示意圖。下面是行為室的示意圖。紅色區域表示鼠標需要進入盒子的區域來觸發容器(藍色)中的食物顆粒的遞送。在獎勵交付后,至少需要30秒的時間才能交付另一個獎勵。b,來自一只老鼠的光度信號示例。上面,行為時間戳是通過紅外光束在容器中測出的。青色,獎賞的容器入口;洋紅色,沒有獎勵的容器入口。中間,動物的中心速度。底部,dLight光度信號。紅色,來自背部的信號;藍色,腹側信號。c,來自示例小鼠的所有試驗的dLight光度信號的熱圖(兩個階段)。上方,來自背部的信號。底部,來自腹側的信號。每一行代表一個單獨的試驗。N = 37次獎勵容器進入試驗。N = 435例無獎勵進入容器的試驗。N = 52次運動啟動試驗。d,小鼠所有會話的平均速度和dLight光度信號。來自背部的信號顯示為紅色;來自腹側的信號顯示為藍色(n = 8次,來自4只小鼠)。陰影區域代表會話平均值的標準誤差。
體內空間分辨光度法
為了更深入地了解構成運動行為和獎賞驅動行為基礎的神經過程,光纖光度法已被用于探測紋狀體神經元的活動11-13,29。在這種情況下,錐形光纖通過使用一個遠程控制的植入物對多個區域進行采樣來擴展實驗能力。為了支持這一論點,我們用紋狀體中的錐形光纖對位點選擇性熒光法進行了表征(圖3f)。我們將一個0.66-NA 錐形光纖插入Thy1-ChR2- EYFP小鼠固定腦片的紋狀體中,在獲得ξ(x,y)場(圖3g)后,我們使用位點選擇性照明來產生增加輸入角度時的光度測量效率ρ(x,y)場(補充圖5)。正如預期的那樣,隨著光輸入角度的增加,響應位點選擇性照明的體積逐漸遠離錐形光纖尖端(圖3h)。
我們使用dLight1.1(參考文獻30)同時測量背側紋狀體和腹側紋狀體的多巴胺瞬變,在體內測試錐形光纖系統,這兩個腦區顯示出不同的多巴胺信號31。我們在一個簡單的操作性條件反射范式中訓練小鼠,在此期間我們從背側和腹側紋狀體收集dLight熒光(圖4a, NA = 0.39)。在這個實驗中,老鼠必須待在房間的一邊,才能觸發食物獎勵從位于房間另一邊的容器中傳遞出來。這迫使老鼠從房間的一邊跑到另一邊去收集獎勵,并在消耗容器中的獎勵時停止移動。
我們在腹側紋狀體中觀察到經典的獎賞驅動的多巴胺瞬變,其中dLight熒光在獎賞受體進入期間增加,在無獎賞進入期間減少(圖4b-d;補充圖6所示的所有試驗均為單獨的小鼠)。然而,對于獎賞和未獎賞的受體條目,背側dLight熒光均下降,這表明背側紋狀體中的多巴胺釋放與運動變化的相關性更強,而不是與獎賞的獲得(圖4b-d)。此外,在獎賞受體進入時,背側紋狀體和腹側紋狀體由行為引起的多巴胺變化的跡象相反,并且與多巴胺在運動啟動中的功能一致32,33,兩個部位的信號在運動啟動期間增加(圖4b-d)。因此,雖然背側紋狀體和腹側紋狀體多巴胺瞬變均追蹤運動,但只有腹側紋狀體的多巴胺瞬變對獎勵有強烈反應。
帶有微結構錐形光纖的設計集光體積
錐形光纖與環境界面的大表面允許根據感興趣的區域設計收集量(圖5)。這可以通過使用微納米制造技術來構建光纖的錐度來實現,正如在光傳輸中所顯示的那樣25,26,34。在這里,我們展示了:(i)光收集在波導周圍特定角度范圍內的限制(圖5a-d), (ii)當光收集被限制在沿錐度的特定點時,在深層皮質層中對細胞體的觀察(圖5e-l), (iii)選擇性照明和從兩個光學窗口收集,通過操縱激發激光束分別解決(圖5m,n)。
為了限制光的收集到波導表面的一半,我們對其對面的高反射鋁層進行熱蒸發(圖5a)。半包被的TF在光學表面的ξ(x,y)(圖5b)和剖面(圖5c)與未包被的雙探針在形狀和收集到的光子數方面相似。因此,金屬涂層并沒有實質性地改變檢測到的信號,這表明幾乎所有進入未涂層錐度的光子都經歷了介電全內反射,這是由半涂層光纖中的金屬層強迫的。我們將半包被的錐形光纖插入由Thy1啟動子控制的表達EYFP的固定腦切片中,在組織中測試了該裝置的側采集特性(圖5c)。我們獲得了一個雙光子的表熒光圖像和ξ(x,y)場,并發現盡管在光纖周圍都產生了熒光,但光纖PMT只獲得了在光纖的未涂覆部分附近產生的熒光(圖5d)。
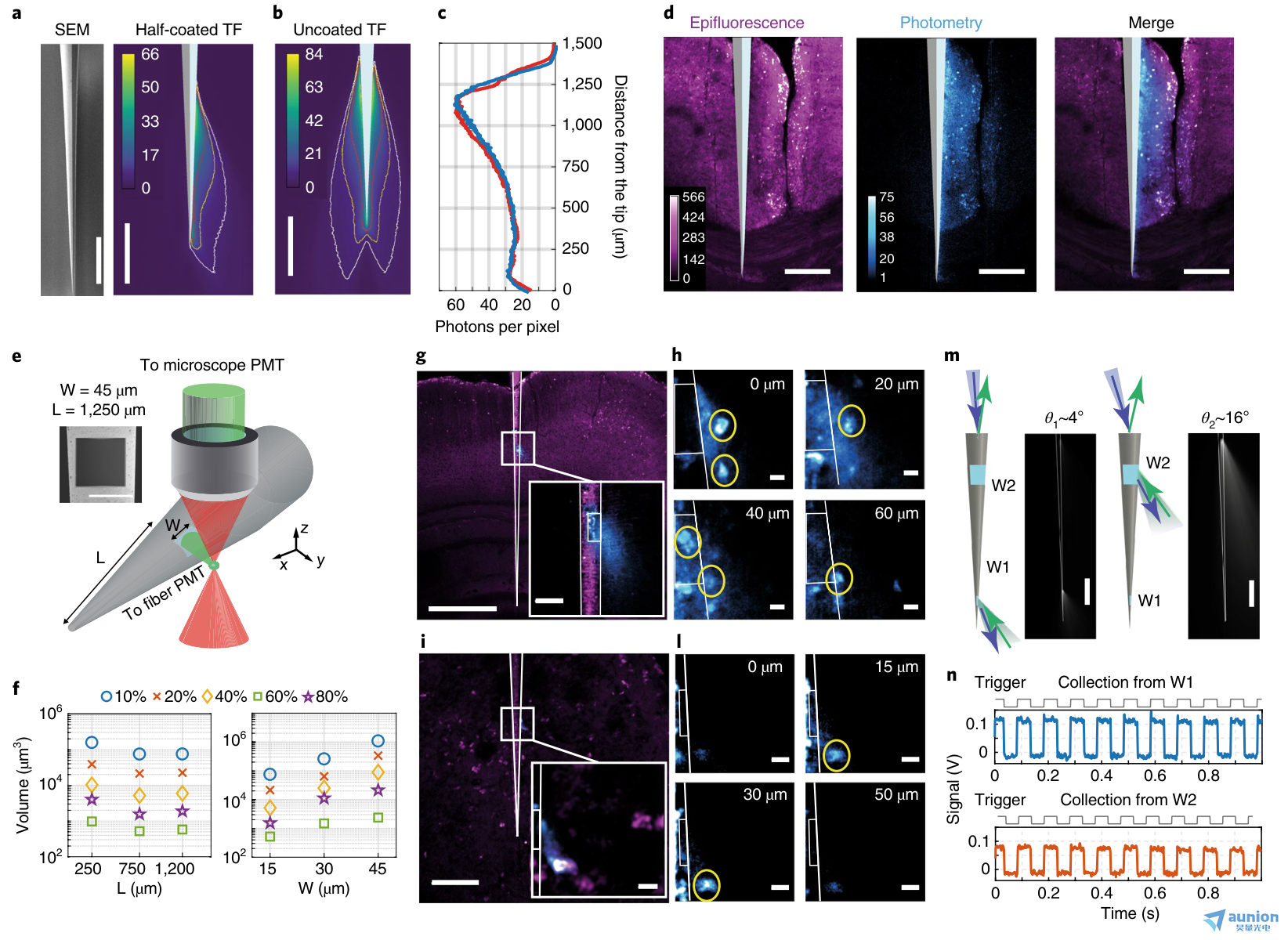
圖5 |用微結構錐形光纖設計集光體積。a,左,半金屬化TF的掃描電子顯微圖(SEM)。右,在pbs -熒光素溶液中,ξ(x,y)場對半涂覆的0.39-NA TF。b, ξ(x,y)場在pbs -熒光素溶液中。等值線分別用白色、黃色和紅色表示每像素5、10和20個光子(停留時間3.2μs)。c, ξ(x,y)對于半涂覆錐形光纖(紅色)和未涂覆錐形光纖(藍色)的場分布。d,在固定的腦切片中收集半涂層TF (Thy1-ChR2-EYFP小鼠)。從左到右:雙光子表熒光,ξ(x,y)場,合并表熒光(品紅)和ξ(x,y)(青色)。比例尺(a-d), 250μm。e,設置用于表征從光學窗口的光收集。插圖,正方形窗的SEM顯微圖(W = ~45μm, L = ~ 1250μm)。f,與L和W相比,由采集等值面包圍的pbs -熒光素溶液的采集量分別為zui大采集量的10%、20%、40%、60%、80%(補充圖6)。g,固定腦片(Thy1-ChR2-EYFP小鼠)的方形窗光采集(W = ~45μm, L = ~750μm);該圖像顯示了ξ(x,y)域(青色)與同時獲得的雙光子表觀熒光圖像(品紅)的疊加;比例尺,500μm。插入,放大窗口;比例尺,50μm。h,在光學窗口上方隨高度增加的Z-stack(0、20、40、60μm)。錐形光纖配置文件和窗口位置用白色表示。比例尺,10μm。i,與g一樣,當TF的時間窗W = ~20μm時,L = ~230μm;比例尺,100μm(插圖,10μm)。l,如h,對于W = ~20μm的光學窗口:高度為0、15、30、50μm的z-stack;比例尺,10μm。m,設置用于選擇地點的照明和收集。錐形光纖被淹沒在pbs -熒光素滴中。激光束脈沖(10 Hz, 50 ms, 473 nm)在受控θ下注入(補充圖7c)。比例尺,500μm。n,從W1和W2窗口測量的熒光信號,通過操縱θ獨立激活(熒光信號通過減去自身熒光校正)。c、d、g-l、n實驗重復3次,結果相似。
從任意深度的光學窗口收集光
我們探索了在全鋁涂層的TF上制造光學窗來進一步限制收集體積的可能性,我們使用聚焦離子束研磨來選擇性地去除錐形特定區域的金屬25,26。為了優化該裝置,我們如上所述對PBS-熒光素溶液(30μM)中光學窗口的光收集進行了表征。我們制作了不同邊長(W = 60、30、15μm)的光學窗平方的探針(NA = 0.39),放置在距離光纖尖端不同距離(L = 230、750、1,250μm)處(圖5e、f)。這些設備的體積收集圖(補充圖7a)顯示,較大的窗口導致較大的收集量(圖5f)。然而,盡管靠近尖端的窗口從略大的體積中收集,但在較大的錐形截面上的窗口具有相似的收集特性(圖5f)。
我們在Thy1-ChR2-EYFP小鼠的固定腦片上,通過采集ξ(x,y)場與同步外熒光成像相關聯,測試了微結構錐形光纖的光學性能。我們發現,對于窗寬W = ~45μm,位于距錐尖L = ~750μm的錐形光纖,光集合與光學窗口位置共定位,其集合葉不垂直于纖維軸,而是指向錐尖(圖5g)。這一特性與來自光學窗的選擇性光傳遞密切相關25,因為它允許在深度上與細胞體積進行界面,具有高空間選擇性(圖5h)。此外,從光學窗口附近的區域獲得的三維熒光堆棧(側W = ~45μm, L = ~750μm)(圖5h和補充圖7b)顯示了光度圖和表觀熒光成像的精確匹配。我們使用窗口寬度W = ~25μm的錐形光纖得到了類似的結果,錐形光纖位于距尖端L = ~230μm處(圖5i, L)。
正如我們所展示的,對于未涂覆的錐形光纖,可以利用分模解復用策略選擇性地激發和收集來自同一錐形光纖不同截面的兩個光學窗口的熒光(圖5m和補充圖7c)。為此,我們將微結構錐形光纖浸入熒光素滴中,并在不同的輸入角度注入473 nm的激光束,以每次選擇一個窗口(補充圖7c)。我們用聲光調制器(10 Hz, 50%占空比方波)調制激光功率時,用光電探測器測量采集到的熒光信號(圖5n)。熒光在每個窗口位置被選擇性激發,表明錐形光纖可以選擇性地照亮和收集來自兩個受限區域的光(圖5m,n和補充圖7c)。
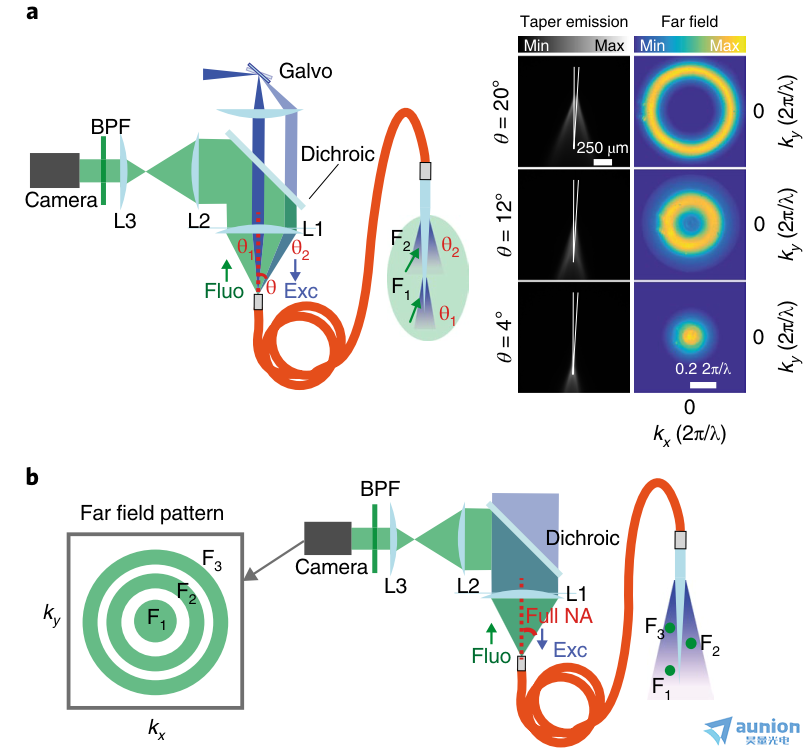
圖6 |利用遠場成像進行深度分辨光纖測光的檢測方案。a、遠場檢測熒光支持時分復用,提高深度選擇性。b,通過全NA刺激實現遠場檢測,實現基于反向傳播熒光的kT值的純分模解復用。Fluo,熒光信號;Exc,激發光。
討論
在神經科學中,從大腦中表達的活動指示器獲取熒光信號是一項強大的技術35,36,可植入式波導系統將極大地造福神經科學領域,該系統可配置為有效和選擇性地收集感興趣區域的光。此外,本文中提出的方法可以在使用遠場檢測來獲得光纖光度實驗中的深度選擇性方面打開進一步的視角(圖6)。
我們設想應用錐形光纖探針從散射組織中收集熒光,將有助于解剖腦深部多個功能區的貢獻,同時為現有的光學方法提供一個通用的補充。基于錐形光纖的分模解復用的光度測定方法也可以潛在地擴展到大類別的軟的、生物的、活組織,其中大腦代表了散射特性方面具有挑戰性的情況之一,考慮到散射長度和各向異性。在這個框架中,錐形光纖為現有的光收集設備增加了有益的功能,使用了扁平切割光纖或μLED/光電探測器系統無法實現的不同配置10。
參考文獻
1. Peterka, D. S., Takahashi, H. & Yuste, R. Imaging voltage in neurons. Neuron 69, 9–21 (2011).
2. Luo, L., Callaway, E. M. & Svoboda, K. Genetic dissection of neural circuits: a decade of progress. Neuron 98, 256–281 (2018).
3. Stroh, A. et al. Making waves: initiation and propagation of corticothalamic Ca2+ waves in vivo. Neuron 77, 1136–1150 (2013).
4. Schwalm, M. et al. Cortex-wide BOLD fMRI activity reflects locally-recorded slow oscillation-associated calcium waves. eLife 6, e27602 (2017).
5. Adelsberger, H., Grienberger, C., Stroh, A. & Konnerth, A. In vivo calcium recordings and channelrhodopsin-2 activation through an optical fiber. Cold Spring Harb. Protoc. 2014, pdb.prot084145 (2014).
6. Fuhrmann, F. et al. Locomotion, theta oscillations, and the speed-correlated firing of hippocampal neurons are controlled by a medial septal glutamatergic circuit. Neuron 86, 1253–1264 (2015).
7. Simone, K., Füzesi, T., Rosenegger, D., Bains, J. & murari, K. Open-source, cost-effective system for low-light in vivo fiber photometry. Neurophotonics 5, 025006 (2018).
8. Muir, J. et al. In vivo fiber photometry reveals signature of future stress susceptibility in nucleus accumbens. Neuropsychopharmacology 43, 255–263 (2018).
9. Adelsberger, H., Zainos, A., Alvarez, M., Romo, R. & Konnerth, A. Local domains of motor cortical activity revealed by fiber-optic calcium recordings in behaving nonhuman primates. Proc. Natl Acad. Sci. USA 111, 463–468 (2014).
10. Lu, L. et al. Wireless optoelectronic photometers for monitoring neuronal dynamics in the deep brain. Proc. Natl Acad. Sci. USA 115, E1374–E1383 (2018).
11. Meng, C. et al. Spectrally Resolved fiber photometry for multi-component analysis of brain circuits. Neuron 98, 707–717.e4 (2018).
12. Cui, G. et al. Concurrent activation of striatal direct and indirect pathways during action initiation. Nature 494, 238–242 (2013).
13. Markowitz, J. E. et al. The striatum organizes 3D behavior via moment- to-moment action selection. Cell 174, 44–58 (2018).
14. Kim, C. K. et al. Simultaneous fast measurement of circuit dynamics at multiple sites across the mammalian brain. Nat. Methods 13, 325–328 (2016).
15. Gunaydin, L. A. et al. Natural neural projection dynamics underlying social behavior. Cell 157, 1535–1551 (2014).
16. Schmid, F. et al. Assessing sensory versus optogenetic network activation by combining (o)fMRI with optical Ca2+ recordings. J. Cereb. Blood Flow. Metab. 36, 1885–1900 (2016).
17. Lütcke, H. et al. Optical recording of neuronal activity with a genetically-encoded calcium indicator in anesthetized and freely moving mice. Front. Neural Circuits 4, 9 (2010).
18. Guo, Q. et al. Multi-channel fiber photometry for population neuronal activity recording. Biomed. Opt. Express 6, 3919 (2015).
19. Cui, G. et al. Deep brain optical measurements of cell type–specific neural activity in behaving mice. Nat. Protoc. 9, 1213–1228 (2014).
20. Tai, D. C. S., Hooks, D. A., Harvey, J. D., Smaill, B. H. & Soeller, C. Illumination and fluorescence collection volumes for fiber optic probes in tissue. J. Biomed. Opt. 12, 034033 (2007).
21. Pisanello, M. et al. The three-dimensional signal collection field for fiber photometry in brain tissue. Front. Neurosci. 13, 82 (2019).
22. Pisanello, F. et al. Dynamic illumination of spatially restricted or large brain volumes via a single tapered optical fiber. Nat. Neurosci. 20, 1180–1188 (2017).
23. Park, S. et al. One-step optogenetics with multifunctional flexible polymer fibers. Nat. Neurosci. 20, 612–619 (2017).
24. Acker, L., Pino, E. N., Boyden, E. S. & Desimone, R. FEF inactivation with improved optogenetic methods. Proc. Natl Acad. Sci. USA 113, E7297 (2016).
25. Pisanello, F. et al. Multipoint-emitting optical fibers for spatially addressable in vivo optogenetics. Neuron 82, 1245–1254 (2014).
26. Pisano, F. et al. Focused ion beam nanomachining of tapered optical fibers for patterned light delivery.Microelectron. Eng. 195, 41–49 (2018).
27. Pisanello, M. et al. Tailoring light delivery for optogenetics by modal demultiplexing in tapered optical fibers. Sci. Rep. 8, 4467 (2018).
28. Pisanello, M. et al. Modal demultiplexing properties of tapered and nanostructured optical fibers for in vivo optogenetic control of neural activity. Biomed. Opt. Express 6, 4014 (2015).
29. Klaus, A. et al. The spatiotemporal organization of the striatum encodes action space. Neuron 95, 1171–1180 (2017).
30. Patriarchi, T. et al. Ultrafast neuronal imaging of dopamine dynamics with designed genetically encoded sensors. Science 360, 1420 (2018).
31. Howe, M. W. & Dombeck, D. A. Rapid signalling in distinct dopaminergic axons during locomotion and reward. Nature 535, 505–510 (2016).
32. Coddington, L. T. & Dudman, J. T. Emergence of reward expectation signals in identified dopamine neurons. Nat. Neurosci. 21, 1563–1573 (2018).
33. da Silva, J. A., Tecuapetla, F., Paix?o, V. & Costa, R. M. Dopamine neuron activity before action initiation gates and invigorates future movements. Nature 554, 244–248 (2018).
34. Rizzo, A. et al. laser micromachining of tapered optical fibers for spatially selective control of neural activity. Microelectron. Eng. 192, 88–95 (2018).
35. Tanese, D. et al. Imaging membrane potential changes from dendritic spines using computer-generated holography. Neurophotonics 4, 031211 (2017).
36. Wang, T. et al. Three-photon imaging of mouse brain structure and function through the intact skull. Nat. Methods 15, 789–792 (2018).
審核編輯 黃宇
-
光纖
+關注
關注
19文章
3950瀏覽量
73514 -
光度測定
+關注
關注
0文章
3瀏覽量
5520
發布評論請先 登錄
相關推薦
光纖線是干嘛用的
光纖怎么看出用幾根光纖跳線
光纖斷了簡易接法
突破傳輸容量瓶頸:多芯光纖與空芯光纖
利用光纖間接檢測 X 射線
室內光纖線用幾芯的

光纖測試的主要方式






 工商網監
工商網監
評論